Definition
Anaphase I is the third stage of meiosis I and follows prophase I and metaphase I. This stage is characterized by the movement of chromosomes to both poles of a meiotic cell via a microtubule network known as the spindle apparatus. This mechanism separates homologous chromosomes into two separate groups.
In anaphase I, the main goal of the spindle apparatus is apparent. This phase can only take place after a positive spindle checkpoint result at the end of the preceding phase, the metaphase. This check make’s sure that the kinetochores – the equivalent of climbing equipment that enables chromosomes to work their way along the microtubule network emitting from the centrioles – are properly attached. Kinetochores are proteins that connect microtubules to the chromosome centromere. Only once all chromosomes are tightly attached to the spindle network can anaphase I begin.
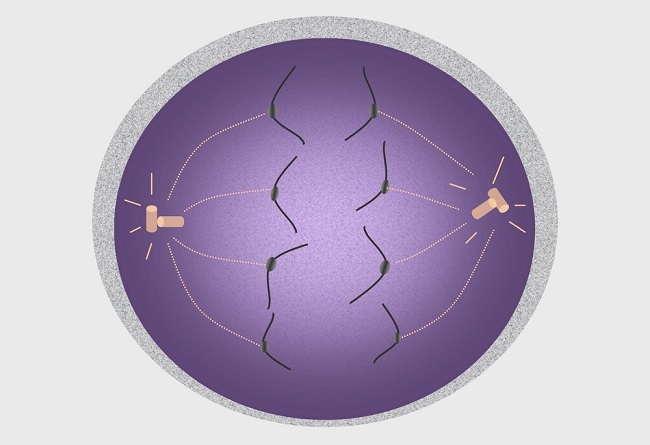
Preparation for Anaphase I
Centrosomes have been commonly believed to be the primary manufacturing units of the spindle apparatus. Centrioles are the main components of the centrosome and appear as short microtubule cylinders in a star-like assembly consisting of groups of three microtubules, as seen in the image below. Centrioles exist in pairs. When combined with pericentriolar material (PCM) they form a centrosome.

A centriole pair is not attached, but during the replication stage (synthesis, or S-stage) of the cell cycle, they are also replicated to form daughter centrioles. The original and daughter centrioles are attached and will remain so until the prophase stage of mitosis or prophase I of meiosis is initiated. It is important that four structures are available, as these will split into two pairs – one pair for each daughter cell.
During prophase, centrosomes separate and migrate to two opposite poles outside of the disintegrating nuclear membrane, where they produce longer microtubules that reach to the cell center (the metaphase plate or equatorial plate). Microtubules have a ‘slow-growing’ (minus) end attached to the PCM of each centrosome, and a ‘fast-growing’ (plus) end which grows towards the metaphase plate.
During prometaphase, kinetochore proteins form close to the centromeres of the chromosomes and attach to the nearest microtubule. If a centromere is damaged, no kinetochore will form and there will be no anaphase. One of each chromosome pair is destined to be carried to one pole, the other to the opposite pole.
During metaphase, each chromosome is aligned and ready to be transported to either pole. As mentioned, a positive result to a spindle check must be completed before the move into anaphase, in which chromosome pairs are separated from their partners and transported to either side of the cell.
What Happens During Anaphase I?
How the chromosomes are brought to each side of the cell during anaphase is now understood as a breakdown of the microtubule network, shortening the microtubule (MT) fibers and hence bringing each chromosome closer to its final destination. Kinetochore function is therefore crucial, as these keep the chromosomes attached, like abseiling rope clips, to a rope that may fray at any moment. As most kinetochores are attached to more than one microtubule – usually a bundle – the breakdown of single sections of microtubule does not lead to the detachment of a chromosome from the spindle, yet this attachment must still be strong. Some scientists liken the kinetochore to microtubule bundle connection to the Chinese finger trap, where any traction forces create an even stronger attachment.
During anaphase I the different microtubule types and functions become important. Kinetochore, astral and interpolar MTs are the three types of microtubule essential for spindle formation. They are formed from layers of alpha and beta tubulin proteins, as shown below.
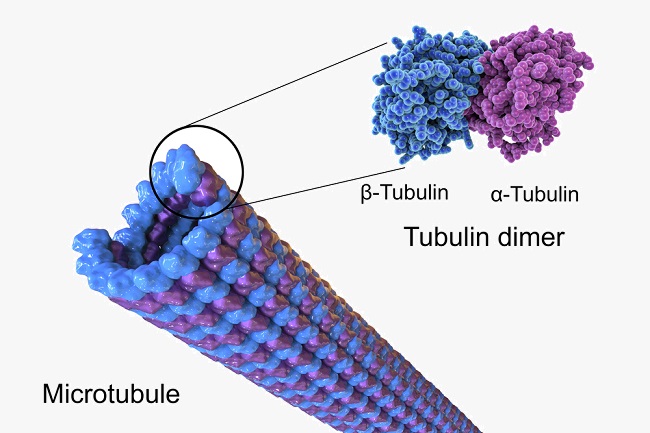
Kinetochore MTs are centrosome fibers connected to chromosome centromeres via a kinetochore. Interpolar MTs are long microtubules that extend past the metaphase plate, overlapping the positive ends of interpolar MTs coming from the opposite pole. Astral MTs do not connect to kinetochores and are thought to create a navigation network for other areas of the spindle network.
Anaphase actually consists of two stages: anaphase A and B. These occur simultaneously but are very different mechanisms. In anaphase A, the connecting fibers of the microtubule spindle shorten through the breaking up of small sections, while kinetochores lead their chromosomes up- or downwards. Electron microscopy usually shows the kinetochore attachment point closer to the pole, with the chromosome arms dangling in the opposite direction. However, some plant and insect studies show the chromosome arms leading, with the kinetochores a little behind. This shows that there are other forces at work in vertebrates which create ‘drag’ on the chromosome arms. These forces are called polar winds. Kinetochores must, therefore, be constantly in motion to prevent a reversal of direction or a to-ing and fro-ing along the spindle, otherwise known as directional instability. This is similar to dynamic instability during anaphase A, caused by the switch between breakage and growth of the microtubules – breakage (shortening) to bring chromosomes closer to their destination, growth (lengthening) to separate the poles even further.
In anaphase B, the spindle network elongates, separating the cell poles even further. Overlapping sections of microtubules – plus ends originating from both poles – slide apart. This mechanism is highly regulated. The cell stretches and elongates, whereby the poles become further apart. This helps to prevent incomplete cell division.
Once a complete set of chromosomes has arrived at either pole of the cell, the next phase – telophase – may begin.
Spindle Apparatus Formation & Organization – Are Centrioles Necessary?
As the organization of the structures necessary for cell division are of great interest to fertility experts, much research has been done concerning spindle construction. An improperly attached chromosome or incompletely developed spindle network during meiosis leads to infertility or miscarriage. The dissolving of the nuclear membrane, allowing centrosomes to migrate to both cell poles, is not timed according to spindle development. This means that the spindle network might not be completed in time for the separation of the chromosome pairs in meiosis I, or of the single chromosomes in meiosis II. Alternatively, early disintegration of the nuclear membrane means that the spindle apparatus can be completely formed prior to anaphase.
Cells that normally contain centrosomes can form spindle networks even after their centrioles have been artificially removed in a laboratory setting. Many plant cells and all oocytes (egg cells) do not contain centrosomes, but microtubule networks still form inside them. It is suggested that the microtubule network is itself responsible for spindle formation, and not the centrosomes. The fact that centrosomes are also responsible for flagellum and cilia formation, and that all male vertebrae cells have centrosomes, might point towards the centrosome as being necessary for flagella, but only complementary for spindle formation. As the ova is a very large gamete, spindle formation from microtubules already within the cytoplasm might be more energy efficient.
However, the large size of the egg can create other problems. At least 10% of human pregnancies produce aneuploid embryos with either extra or missing chromosomes. The most commonly quoted of these is trisomy 21, or Down syndrome. This is because atypical spindle forming and positioning can lead to asymmetrical cell division and the loss or gain of one or more chromosomes to either daughter cell.
The image below shows the large difference in size between egg ova and spermatozoa, and the long flagellum of the male gamete. The presence of centrioles in the male gamete and not the female may therefore be more important for flagellum formation than the spindle network.

Separating the Tetrad
Due to replication during a precursory phase in mitosis (non-gamete cell division) the chromosome pairs separated during anaphase I are different from the chromosome pairs in the normal human karyotype.
Before preparation for either mitosis (somatic cells) or meiosis (gametes or sex-cells) a human cell contains 46 loosely wrapped chromatids within its nucleus. These are arranged in pairs – one from the mother and one from the father. This means the non-dividing human cell contains 23 pairs of chromatids, the total sum of which is 46 chromatids. The sole difference between a chromatid and a chromosome is the packaging – when loosely packaged the complex of DNA and binding proteins is called a chromatid, when tightly packaged it is called a chromosome.
A tetrad (or bivalent) is specific to the process of meiosis. It is the result of replication in the S-phase of the natural cell cycle in combination with the meiosis-specific procedure of crossing over, the result of which is recombination – the mixing up of alleles across a chromosome pair. Upon recombination, a pair of chromosomes is referred to as a tetrad. In mitosis, crossing over does not occur and the resulting replicated chromosome pairs are just that, chromosome pairs. So the tetrad refers solely to a pair of recombined chromosomes during meiosis.
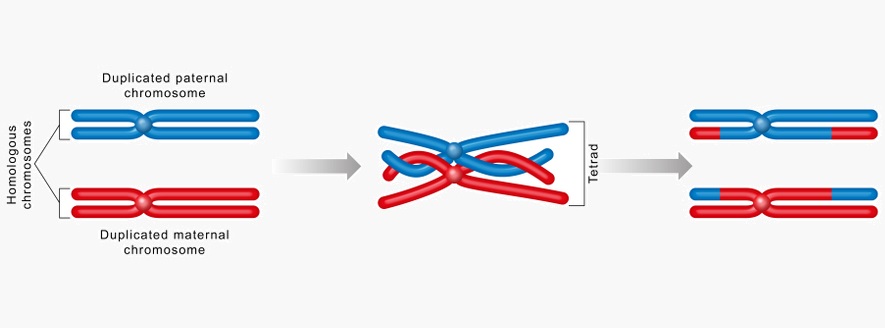
Because tetrads are not exact copies, their development and eventual sharing during reproduction lead to genetic variation across a species. In order to divide, a cell requires a complete set of genetic material in the form of 23 chromatid pairs. Yet meiosis is a two-stage cell division where a single cell forms four daughter cells, each with a single chromosome. In the first stage – meiosis I – a single cell divides into two daughter cells, each containing 23 pairs of chromosomes or 46 chromosomes. In meiosis II, these two daughter cells split again, this time each containing 23 chromosomes, not chromosome pairs. As the only cells that are created by meiosis are sperm and egg cells, this makes sense. During fertilization, a single zygote is formed from two gametes which must contain 23 chromosome pairs – a combination of the genes of both mother and father.
Is Anaphase I of Meiosis and Mitosis Different?
During a phase of the cell cycle called the S-phase (synthesis phase), the DNA that is the main component of the chromatid is copied or replicated. This creates two near-exact strands for each of the 46 chromosomes. To make sure they remain close to each other and do not cause genetic defects through ‘lost’ genetic material, these sister chromatids are joined by a centromere.
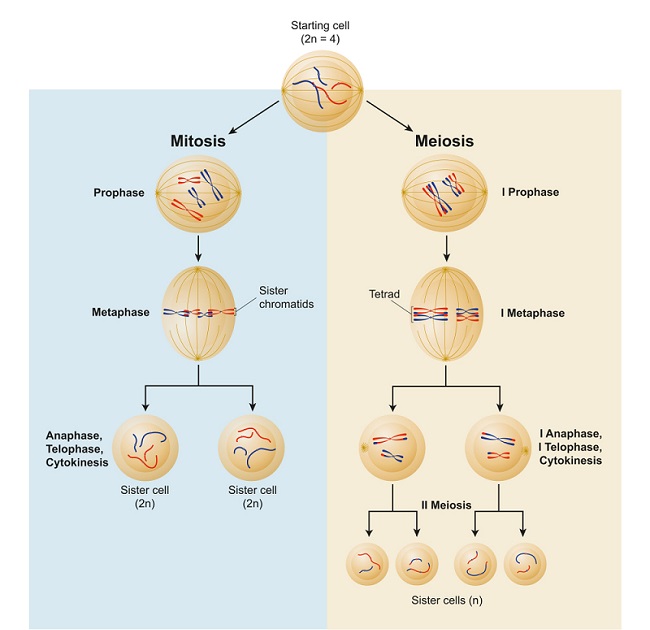
Firstly, there is no anaphase I in mitosis, only anaphase. Mitosis is a single-step process where one cell becomes two. Meiosis is a two-step process, first creating two cells out of one, and then four cells out of those two.
While DNA replication is the same in both mitosis and meiosis during the normal cell cycle, once a cell moves into either division process, the differences begin. This is because the goals of mitosis and meiosis are completely different. The aim of mitosis is to create an as exact as possible copy of a cell from a single source. Meiosis, on the other hand, wants a mix of genetic information that can be passed over to the next generation from two completely different sources (mother and father). Some of the differences between these two processes can be seen below.
Explaining the Difference Between Mitosis and Meiosis – the Stripey Sock Analogy
To make it easier, imagine two non-matching striped socks. One is striped black, white and yellow. The other is striped red, blue and green. They represent a single chromosome (or chromatid) within a non-dividing cell. There are 23 chromosome pairs in a single human cell, but imagining 23 different colored pairs of socks is quite confusing, so it is easier to stick to a single chromosome (or one pair of stripey socks).
When replication occurs (before either mitosis or meiosis) and the cell enters the S-phase, these 2 socks are each given an exact twin. A total of four socks. The two black, white and yellow socks are clipped together (clip = centromere), and the two red, blue and green socks are, also. They now represent a replicated chromosome pair.
At the conclusion of mitosis, a fat cell divides to produce two fat cells, or a hepatocyte divides to make two hepatocytes, and so on. It is therefore important that this genetic information remains the same. The two pairs of socks representing the chromosome are separated. The clips are pulled off and one set of socks goes to one cell, and the other set goes to the other. Two exact copies of the original.
In meiosis, the outcome will be a new human and a combination of two completely different people. However, the replication method is the same, and there are two black, white and yellow socks clipped together, and two red, blue and green socks clipped together. During meiosis, an exact copy is not the desired outcome, so the red of one sock swaps with the white of one of the other colored socks (crossing over). We now have one black, white and yellow sock, one red, blue and green sock, one black, red and yellow sock, and one white, blue and green sock. They remain clipped together, but are no longer exact pairs – they have undergone recombination and therefore represent a tetrad – two completely different sock pairs. The pairs are now ready to be pulled to different sides of the cell during anaphase I. This means that the resulting two daughter cells both contain a slightly different but complete set of genetic information – two clipped-together socks each. Of course, this is extremely oversimplified, as hundreds of alleles will be swapped between chromosomes. But this analogy is meant to keep a confusing subject simple to understand.
Quiz