Definition
During prophase II of meiosis II, four important steps occur. These are the condensing of chromatin into chromosomes, disintegration of the nuclear envelope, migration of centrosomes to either pole, and the reconstruction of the spindle apparatus. However, centrosomes are not present in all cells.
What Happens During Prophase II?
Meiosis II occurs in both daughter cells that were formed during meiosis I. As no DNA replication takes place in this second step of meiosis, the cell division process immediately begins. Prophase II prepares the cell for secondary meiotic division where two haploid cells eventually form four haploid cells, each containing half of the genetic information previously contained in the original, replicated diploid cell. A much less complex phase than prophase I, prophase II does not include the steps of leptotene, zygotene, pachytene, diplotene, and diakinesis but more resembles the simpler process of mitotic prophase in non-gamete (somatic) cell division.
Prophase II begins immediately after cytokinesis – the splitting of the diploid gamete into two haploid daughter cells. In order to prepare for the next division, prophase II condenses chromatin first into chromatids, and then more tightly into chromosomes. Simultaneously, the nuclear membrane dissolves, leaving an open area of cytoplasm in which a network of proteins (microtubules) has enough space to create pathways reaching from one side of the cell to the other – the spindle apparatus.
After the condensing of chromatin into chromosomes, and after the disintegration of the nuclear envelope containing these chromosomes, the centrosomes migrate to either pole. Even in the absence of centrosomes, the reconstruction of the spindle apparatus used during meiosis I can be initiated. These four steps characterize prophase II.
Building a Spindle Apparatus Without Centrosomes
Current research is looking at the role of centrioles in human spindle apparatus formation, as female gametes – oocytes – do not have them. Centrioles are constructed from microtubules and, certainly in male gametes and during mitotic cell division, play an important role in spindle apparatus construction. During the normal cell cycle, centrioles replicate to form pairs that are later enveloped in pericentriolar material (PCM). From this point on they are referred to as centrosomes. Two centrosomes, each containing a centriole pair, migrate to either end of the cell – the poles. During mitosis and in male gametes, centrosomes are responsible for constructing a network of microtubules which extend from within the centrosome towards the cell’s center. Construction of this network occurs at a later phase.
However, female reproductive cells have been discovered not to contain centrosomes and instead form the spindle apparatus from existing microtubules within the cytoplasm. The lack of a chromosome or presence of an extra chromosome in daughter cells caused by faults during chromosome separation is known as nondisjunction, where resulting gametes produce abnormal embryos. Nondisjunction of chromosomes is the primary cause of infertility and miscarriage, as it is during female gamete division (meiosis I and II) that chromosomes are most likely to be unevenly divided between daughter cells. Even then, most sources list centrosomes as the manufacturers of the microtubules which make up the spindle apparatus. This may be true in terms of spermatozoa and somatic cells, but does not apply to higher plants and the ova of many other species, including the human race.
This means that, while it is understood that most human cells require at least two centrioles, the fertilized zygote contains only one which it inherited from the male gamete. As with DNA, male gamete centrioles are not replicated during prophase II. Yet, as the zygote grows via mitosis, further cells do contain two centrioles; it is not yet fully understood how the second centriole is formed but its appearance may be linked to precursor proteins contained either in the spermatozoa or ova.
Difference between Prophase I and II
Prophase I consists of five stages. These five stages are not represented in prophase II. Prophase II encompasses four different mechanisms; namely the tight wrapping of DNA into chromosomes, the dissolving of the nuclear membrane, migration of the centrosomes (when present), and the reconstruction of the spindle apparatus.
In prophase I, the first stage is known as leptotene. This stage involves the unwinding of the DNA structure to enable an exchange of alleles between homologous chromosome pairs. No crossing over occurs in prophase II. Therefore, prophase II does not feature leptotene.
The second stage of prophase I is called zygotene. This involves the attachment of a chromosome pair prior to crossing over. As no crossing over occurs in prophase II, this stage is also not included. This is also the case for subsequent phases: pachytene (crossing over), diplotene (breaking down of the crossing-over network), and diakinesis (movement of crossing-over channel to chromatid arms).
However, some parts of prophase I and II are similar. These include the dissolution of the nuclear envelope and centrosome migration. Even so, prophase II of meiosis II is usually compared to the prophase of mitosis, where nuclear envelope breakdown occurs together with centrosome migration, condensation of chromosomes, and the formation of the spindle apparatus. In prophase II, the correct term for the latter of these mechanisms is reformation, as the microtubules that made up the spindle apparatus previously constructed in meiosis I are still available.
And Why the Difference?
When looking at the differences between primary meiotic division and secondary meiotic division, it is always helpful to remember the goal of meiosis – to produce four different gametes (spermatozoa or ova) containing a single but complete portion of genetic data which, when combined with another gamete, creates a cell (zygote) which has a complete (double) set of genetic information from each of two parents.
Genetic Variation
To ensure that this genetic information is not exactly the same and thus contributes to the genetic variation within a species, meiosis I includes a phase in which alleles are swapped between a chromosome pair (crossing over) to produce recombinant chromosomes. This must occur before the cell divides, and takes place in the first prophase of meiosis I. Once this has occurred, the two daughter cells will contain slightly different genes.
This means that there is no need for crossing over during the second cell division. Also, as crossing over only occurs between two replicated, paired chromosomes this is simply not possible in a haploid cell, which does not contain replicated pairs, just pairs.
The distinction between chromosome pairing and replication is often confusing, as when one considers the second cell division in meiosis II, some images seem to show a single X-shaped chromosome being split in half to provide half a chromosome. This is very incorrect.
The human chromosome, as seen on the karyotype below, consists of 46 single chromosomes. Human DNA holds the genetic data for the entire human body within these 46 chromosomes, yet this data is repeated as it is sourced from two parents. Using chromosome 1 as an example, which contains approximately 8% of the genetic information required to produce a human being, we can iron out a lot of unnecessary confusion.
In the image below, two strands of DNA make up chromosome 1. These single strands are usually referred to as chromatids, although this is technically incorrect as the difference between chromatid and chromosome is more to do with the way in which the DNA molecule is packaged. However, by calling each strand a chromatid, the replication process becomes less confusing.
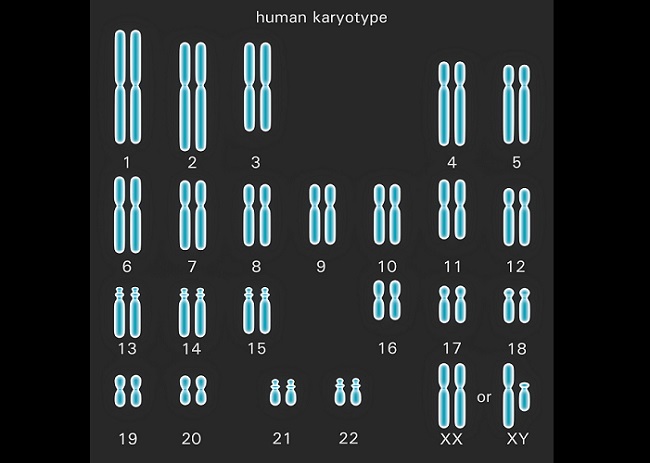
One chromatid derives from the spermatozoon of the father – a full set of data in the cell created during the final stages of meiosis II. The other chromatid derives from the ovum of the mother. Again, a full set of data in each somatic cell similarly created during the final stages of meiosis II. Together, both chromatids contain two sets of information with slight differences – they have the same genes at the same positions (loci), but may contain different alleles.
Lack of a Homologous Chromosome Pair
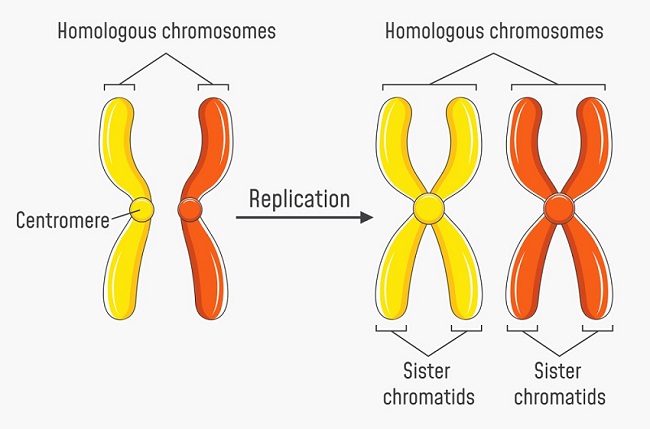
The default number of chromatids in any human cell (apart from the gamete) is 46. During preparations for cell division these 46 chromatids pair up – like to like. Chromatid 1 of the father draws close to chromatid 1 of the mother, and so on. In this example, homologous chromosome 1 is the result. Throughout the human karyotype, 23 pairs of homologous chromosomes which are not physically attached to each other are the result. A cell which contains 23 pairs of homologous chromosomes is known as a diploid cell. Below, the left-hand image shows a pair of non-replicated homologous chromosomes (chromatids); yellow and orange denote the genetic information provided by each parent.
During the replication process of the cell cycle and prior to any type of cell division, all DNA is replicated. Chromatid 1 of the father is replicated, as is chromatid 1 of the mother, and so on. Each replicated chromosome is attached to its copy by way of a centromere, forming the typical X-shape often seen in textbooks. Chromosome 1 no longer consists of two separate chromatids, but of a pair of twin or ‘sister’ chromatids. The complete human karyotype still consists of 23 pairs of chromosomes (one from the father, one from the mother), but each parent’s single chromatid strand has been doubled to make two sister chromatids. Whereas before replication the human karyotype is represented by 23 homologous pairs of 46 chromatids, the replicated karyotype is represented by 23 homologous pairs of 92 chromatids. It is therefore always important to either indicate or be aware of whether a chromosome pair is replicated or not. The right-hand image below clearly shows the original and replicated sister chromosomes in a single homologous chromosome pair.
In the meiotic prophase I, crossing over occurs. This process swaps over a variety of alleles in order to produce sex cells (gametes) which are not clones of either parent. Once crossing over has occurred the homologous pair is known by other names – the tetrad, the bivalent or recombinant chromosomes/chromatids. It is easy to see that crossing over occurs between similar alleles of each parent’s chromatids in the diagram below. Genetic data is not exchanged between sister chromatids of a single chromosome but between the chromatids of the homologous pair. The absence of homologous pairs in haploid cells is the reason why no further crossing over occurs during prophase II.
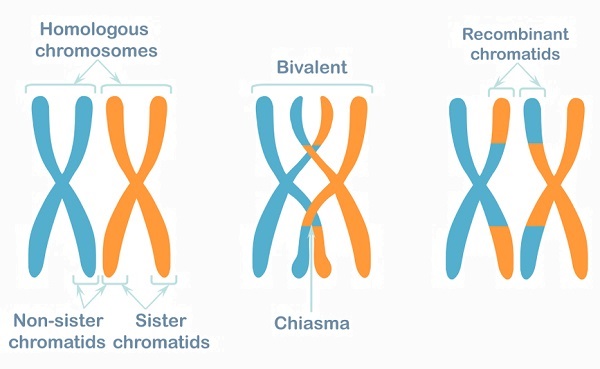
After crossing over, the tetrads (recombinant chromosome pairs) can be separated. Tetrads contain 23 chromosome pairs composed of 92 chromatids. Over the next stages of meiosis I, each tetrad is separated from its paired partner (and is therefore no longer called a tetrad, bivalent or recombinant pair). Instead, a single replicated chromatid (X-form) moves to one side of the cell, and the other half of the former pair moves to the other side. As recombinant replicated chromatids, they contain a mix of alleles from both parents.
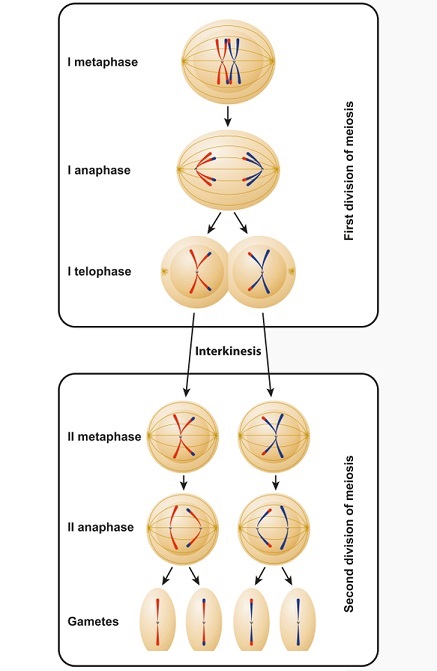
In prophase II, no crossing over occurs as this must take place between homologous chromosome pairs. The spindle apparatus forms in order to break apart the replicated chromatids. In a female, this results in secondary polar bodies, each containing a single chromatid (it can just as easily be called a chromosome) which itself contains the entire human genome. One or more of these will develop into an ovum. In males, the result is four spermatozoa. The difference is clear in the image below, with red and blue chromosomes or chromatids depicting the parentage, although after metaphase I small parts of genes are crossed over.
Two Sets of Chromosomes From Two Different Sources
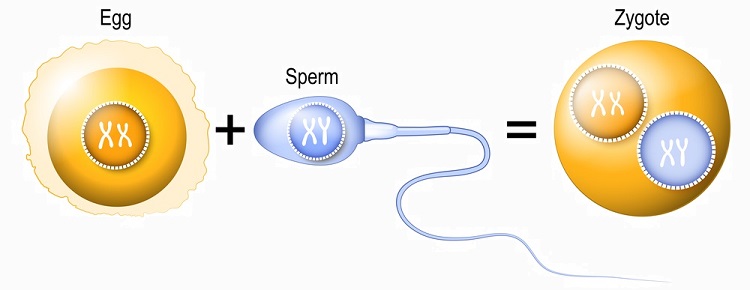
The goal of meiosis is to produce offspring of the same species but with genetic variations. The following image shows recombinant chromatids in egg and sperm combining upon fertilization of the egg to produce a zygote. The zygote is described as a haploid cell as it contains chromosome pairs but these are separated by nuclei. Only during its first mitotic division will these nuclei dissolve and allow the chromosomes of both parents to align on the spindle apparatus as pairs. The resulting two embryonic cells will be diploid.
In the example below, sperm meets egg to produce a zygote containing both sets of genetic information. However, this image is a typical example of why replication and chromosome pairs are often confusing or misinterpreted. The genetic information contained within egg, sperm, and zygote is depicted here as replicated pairs. Immediately upon fertilization, egg, sperm, and zygote should not feature X-shaped chromosomes but single-strand chromatids. Only in preparation for the zygote’s first division, and after DNA replication has taken place, can this genetic information be correctly represented by X-shaped chromosomes.
Quiz