Definition
The first metaphase of meisosis I encompasses the alignment of paired chromosomes along the center (metaphase plate) of a cell, ensuring that two complete copies of chromosomes are present in the resulting two daughter cells of meiosis I. Metaphase I follows prophase I and precedes anaphase I.
Metaphase I Explained
In meiosis I, the lining-up stage of metaphase I is relatively rapid. Tetrads or bivalents (a pair of chromosomes with four chromatids (2 originals, 2 copies) are pulled into line at what is known as the metaphase (or equatorial) plate. This plate does not actually exist but is an imaginary central line along which the chromosomes are positioned.
The individual chromosome in each pair remains close to its partner and lines up one on top of the other. This will eventually lead to one chromosome migrating to one pole, the other to the opposite pole. It also does not matter in which direction these chromosomes are horizontally oriented along this imaginary line. The DNA from either parent can face either side of the cell. This increases gene variation, as where one daughter cell might contain 40% of the father’s chromosomes and 60% of the mother’s, the other will have 60% and 40% of each parent respectively. This would account, for example, for a first child having the father’s eyes and the mother’s nose, and the second having the mother’s eyes and the father’s nose. Due to recombination in the prophase, either child will not look exactly the same as either parent, only similar.
In the image below of an onion cell (in metaphase I), the dark-purple stained chromosome pairs are all centrally positioned along the metaphase plate.
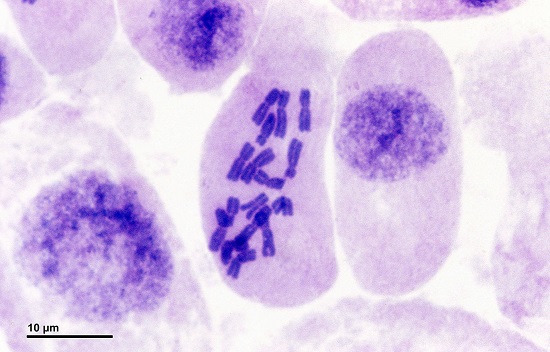
In metaphase I, the two chromosomes of a homologous pair face opposite poles. As recombination has taken place, each of the four chromatids (and, of course, both homologous pairs) have slightly different genetic material. Further steps will pull one of the homologous pair to one end of the cell, and the other to the opposite end. This means that the resulting two cells (produced during cytokinesis at the end of meiosis I) will contain full sets of chromosomes, but these will not consist of chromosome pairs.
The Confusion Surrounding Chromosomes and Chromatids
This allocation of chromosomes is particularly important to understand but often leads to much confusion. It is worth looking at this topic from a fresh perspective, taking a few steps back in the process.
The human karyotype is composed of 22 chromosome pairs and one pair of sex chromosomes (XX or XY). A total of 23 chromosome pairs. Every non-gamete cell contains this set of 23 pairs within its nucleus, except for mature red blood cells or old cornified cells. The two chromosomes that make up a homologous pair come from an organism’s parents – one from the father and one from the mother. In the karyotype below of a male human, these pairs are very distinct. Note the presence of 23 pairs, and a total of 46 chromosomes.
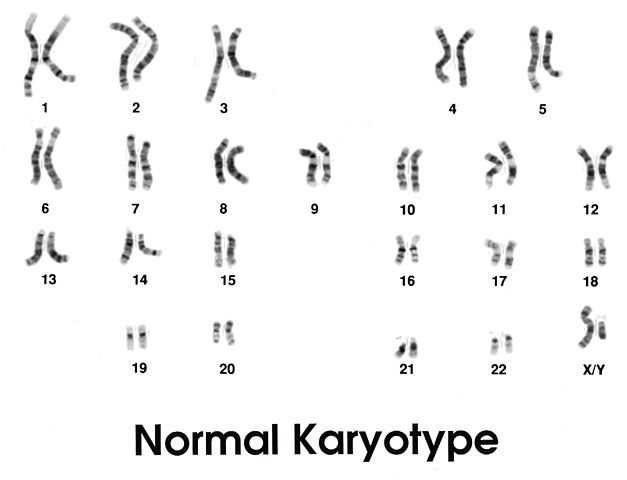
In this image, no X-shape can be seen. Each chromosome is a single strand. Each single strand is a chromatid with a centromere (not visible). This is because the X-shape exists only after the replication of DNA occurs during the S stage of the interphase which precedes both mitosis and meiosis.
The great confusion many students encounter when studying meiosis is the traditional idea that chromosomes are X-shaped. However, this is only the case after each chromatid has been replicated to create two sister chromatids joined together at the centromere. Before replication, a human set of 23 chromosome pairs (each pair consisting of one chromosome from the father, one from the mother) has 46 chromosomes, as we can count in the above image. Each of these 46 chromosomes consists of a single chromatid. The human karyotype before replication, therefore, has 46 chromosomes (23 pairs) and 46 chromatids.
After the S stage of interphase, the chromatids are replicated and the copies remain attached to the ‘sister’ via the centromere. The final count is still 23 chromosome pairs, but each chromosome now consists of two sister chromatids. This extra chromatid provides the X-shape of the chromosome as we are used to seeing it. There are still 46 chromosomes (23 pairs), but now there are 92 chromatids. As the combination of two chromosomes (a pair) containing 4 chromatids (two original, two replicated) is also known as either a tetrad or a bivalent, we can also say that a tetrad has two chromosomes and four chromatids. Tetrás in Greek means four, while bi relates to two.
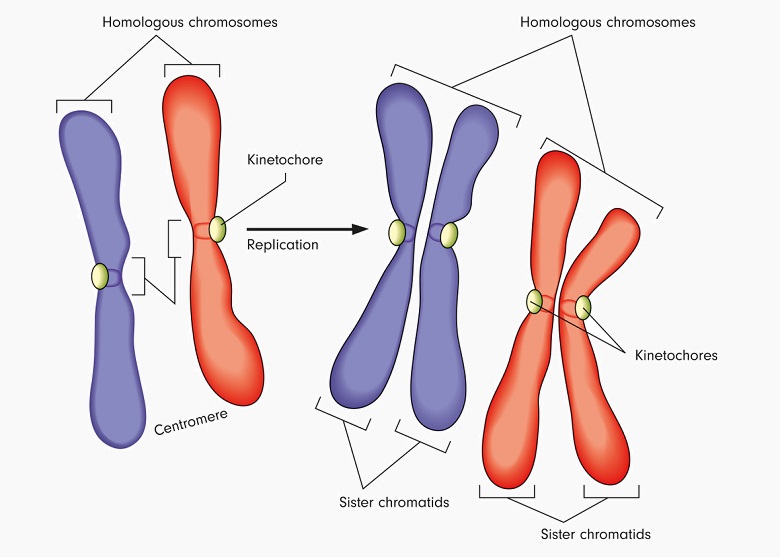
This all becomes much clearer when we think of the goal of meiosis and the subsequent fertilization of an egg by a sperm. Meiosis is a two-step process (with many sub-steps) that first splits a single cell into two cells, each with a complete but slightly different set of DNA contained in chromosome pairs. This is why DNA must be replicated beforehand. If not, it would not be possible to provide a full set of chromosome pairs to the two daughter cells. The end result is two daughter cells, each containing 46 chromosomes (23 pairs) and 46 chromatids.
These two cells then split a second time under meiosis II, creating four daughter cells which each contain a complete set of DNA which is not presented in chromosome pairs, but as a single chromosome. Each cell now contains 23 chromosomes and 23 chromatids. These cells are produced as sperm in males and eggs in females. Upon fertilization, the single chromosomes of each sex combine, once again creating a full chromosome pair.
Chromosome Alignment Along the Metaphase Plate
The imaginary line through the center of a cell called the metaphase plate requires the spindle apparatus in order to align the matching chromosome pairs along its length.
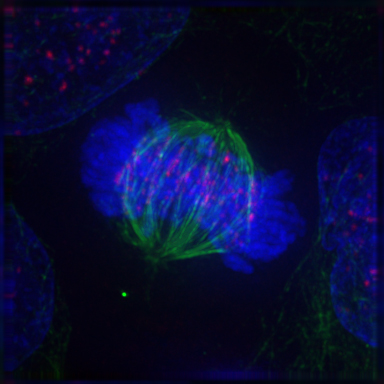
Centrioles, protein structures that send out fibers known as microtubules, position themselves at opposite sides of the cell, creating two poles. Long protein fibers called microtubules are then sent out from both centrioles, forming the spindle apparatus. These microtubules meet (but do not join together) at the center of the cell where the chromosome pairs line up. Once the spindle apparatus is constructed, an attachment occurs between each chromosome’s kinetochore, found close its centromere. This attachment is very stable and has been compared to a Chinese finger trap – the higher the tension, the stronger the attachment.
If we imagine the centrioles lie at the north and south of a circular cell, with the chromosome pairs lined up through the equivalent of the equator – one of the pair above the equator and the other below – it is easy to also imagine how these chromosome pairs can be separated, one pulled up, the other down along these fibers. This migration does not happen during the metaphase. Instead, microtubules pull at the chromosomes until they are properly aligned. The cell then checks the chromosomes are correctly positioned before moving on to the next phase of meiosis I.
Quick Overview of Meiosis I
The metaphase is a single, short stage of the long process of meiosis and occurs twice – once during meiosis I and once during meiosis II. In order to place this particular stage within the meiosis I process, it is important to be at least slightly familiar with the stages or phases that come before or after the metaphase.
Meiosis I begins with the interphase, moving on through the stages of prophase I, prometaphase, metaphase I, anaphase I, and finally telophase I and cytokinesis, and is a procedure in which homologous chromosomes become separated. Meiosis always produces sex cells or gametes, in the form of eggs or sperm.
Interphase
Interphase consists of four stages, the first of which – Gap 1 – is the stage where most eukaryote cells spend the majority of their life and is characterized by protein synthesis which promotes cell growth, function, and health. The interphase occurs in cells before both the process of mitosis and meiosis. At this phase, DNA is not tightly bound in chromosome form, but loosely bound as chromatin. As chromatin, DNA replication is possible. Tightly bound as a chromosome, it is very difficult to replicate.
The second stage – S or ‘synthesis’ – describes DNA copying in preparation for cell division prior to either mitosis or meiosis, where protein histones are produced to wrap the newly copied DNA strands. This phase also includes a higher production of phospholipids which are used to form the membranes of the new cells.
Stage 3 – Gap 2 – describes the replication of cell organelles, and once again protein synthesis is crucial. At this stage, it is still possible to repair faults in the DNA.
Another stage – Gap 0 – is not always the final stage of the interphase. It can occur prior to G1, during G1 or after cell division. Cells that enter G0 can do so temporarily and go on to divide, or leave the replication cycle completely and enter the G0 process which turns them into terminally differentiated cells.
Prophase
Prophase I is characterized by DNA condensation into chromosomes, synapsis, crossing over with resulting recombination of alleles, the migration of chiasmata to the ends of the chromatid arms, and the movement of these arms towards different poles, pulling the chromosomes very slightly apart. The nuclear membrane also starts to dissolve in preparation for the migration of microtubules which form the spindle apparatus.
Prometaphase
Often forgotten, the prometaphase is synonymous with the completion of the breakdown of the nuclear envelope or membrane and the formation of kinetochores. Kinetochores form at a ratio of one per chromosome so that the first movements of the chromosomes to opposite poles using the spindle apparatus can begin. The prometaphase is often relegated to the final stage of prophase I. Kinetochores are proteins that create fixation points on the chromosomes whereby the chromosomes are able to attach themselves to the spindle apparatus and move along it. This might be compared to the carabiners or climbing clips of mountain climbers or abseilers which connect the climber to the rope, allowing him or her to ascend and descend.
Metaphase
The metaphase, into which this article has gone into much more detail, is also known as the ‘lining up’ stage. Spindle fibers from centrosomes at both cell poles connect to the tetrad kinetochores, lining them up through the middle of the cell.
Anaphase
During anaphase, cells grow in size. This growth pulls the opposite poles even further apart and provides traction forces that can eventually pull each chromosome apart and to opposite ends (poles) of the cell.
Telophase
Once the chromosomes reach their opposite poles, the telophase begins. The spindle apparatus breaks up, while two nuclear envelopes form around the two sets of chromosomes, which start to unravel into ‘string of beads’ chromatin.
Cytokinesis
Cytokinesis is the actual division of the cell. A groove (called a furrow) develops in the middle of the cell and narrows while the microtubules in the cytoplasm contract, pinching a single diploid cell into two haploid daughter cells. In plant cells, the cell membrane separating the two cells is replaced by a cell plate.
Quiz