Fluid Mosaic Model Definition
The fluid mosaic model is one way of understanding biological membranes, consistent with most experimental observations. This model states that the components of a membrane such as proteins or glycolipids, form a mobile mosaic in the fluid-like environment created by a sea of phospholipids. There are restrictions to lateral movements, and subdomains within the membrane have specific functions.
Development of the Fluid Mosaic Model
This model was developed over many years, through painstaking work of scientists across the world. It began with the hypothesis that the membrane was made of a lipid bilayer, where membrane phospholipid self-assembled into a dual layer, with the non-polar, hydrophobic tails facing each other. The hydrophilic ‘head’ regions face the cytosol and the extracellular region. This was verified by extracting cell membranes and spreading the lipids into a single layer. The overall surface area of this monolayer was twice that of the plasma membrane, supporting the idea that the lipids formed a bilayer.
However, this was only the beginning, since it became apparent that cell membranes had to be made of more components, to account for their varied biophysical properties. For instance, unlike pure lipids, cell membranes don’t freeze very easily. The permeability of the membrane to bulky polar molecules could also not be explained.
More than 25 years after the lipid bilayer model was proposed, cell membranes were first visualized in the 1950s. The initial observations seemed to suggest that the lipid membrane was coated on either side by thin sheets of proteins. However, in 1972, two scientists, Singer and Nicolson, refined this to create the fluid mosaic model. In this, the phospholipid bilayer was said to be punctuated by various proteins that formed a mosaic-like pattern in the lipid membrane. These proteins could traverse the entire membrane, or interact with one of the two lipid layers. Some proteins could even be attached to the membrane only through a short lipid chain.
The membrane is fluid, but also has an underlying structure, being anchored to the cytoskeleton. The fluid nature of the lipid matrix forming the membrane was first illustrated by an experiment where membranes with different compositions were artificially fused. The proteins of both cells redistributed themselves across the entire fused membrane in less than an hour.
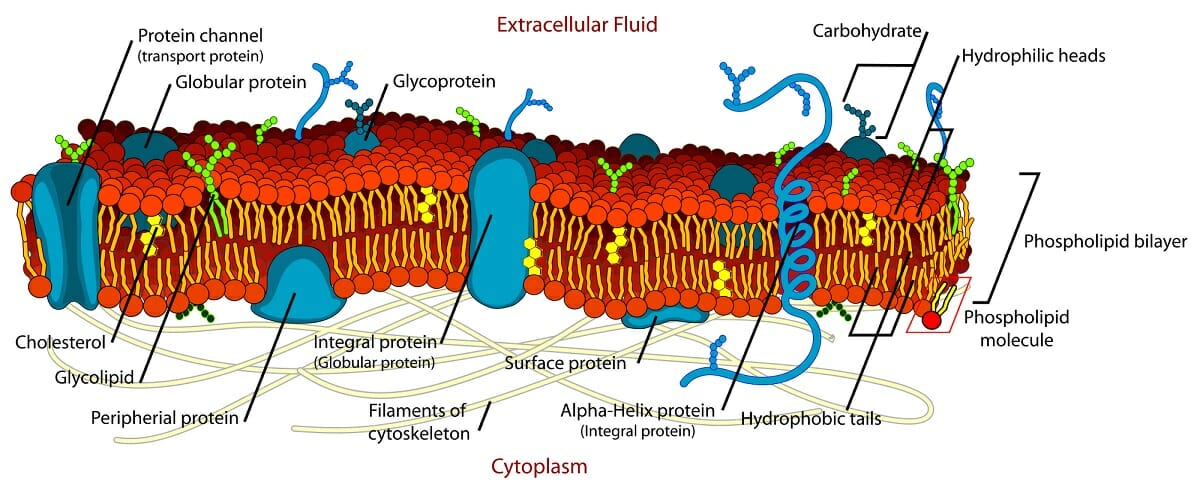
The image illustrates this model, and depicts the lipid bilayer, with different types of integral membrane proteins, as well as cholesterol, glycoproteins, and glycolipids. The image also shows the anchoring of the membrane to the cytoskeleton.
Now, with advanced imaging techniques, some membranes have been studied in great depth, to a resolution of less than a nanometer. These images can even reveal the relative location of different polypeptide chains and lipids within the membrane.
Functions and Components of Biological Membranes
The main function of cell membranes is to demarcate the inner and outer regions of the cell. Within the cell, membranes of organelles perform the same function for subcellular structures.
This function comes along with a caveat – the cell needs to actively communicate with the external environment, exchange materials, while also retaining important nutrients and keeping harmful substances out. The components and structure of biological membranes help in fulfilling these roles and maintaining their selective permeability.
Biological membranes, especially cell membranes are made of phospholipids, cholesterol and proteins.
Phospholipids
The first is the phospholipid bilayer itself that creates a hydrophobic layer separating aqueous environments on either side. Phospholipids are amphipathic molecules consisting of a polar, hydrophilic ‘head’ region formed by a phosphate group and the non-polar hydrophobic ‘tail’ consisting of two long chain fatty acids. These two segments are covalently attached to a glycerol molecule.
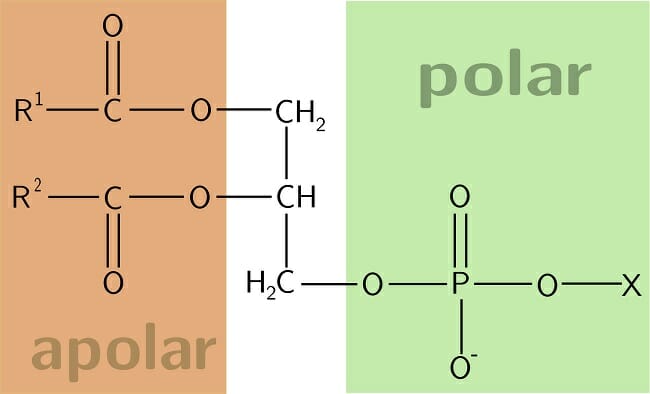
Image shows a schematic representation of the chemical structure of a phospholipid, with R1 and R2 referring to the two fatty acid chains. Usually, one of the two fatty acids will be unsaturated, with at least one double bond between two carbon atoms.
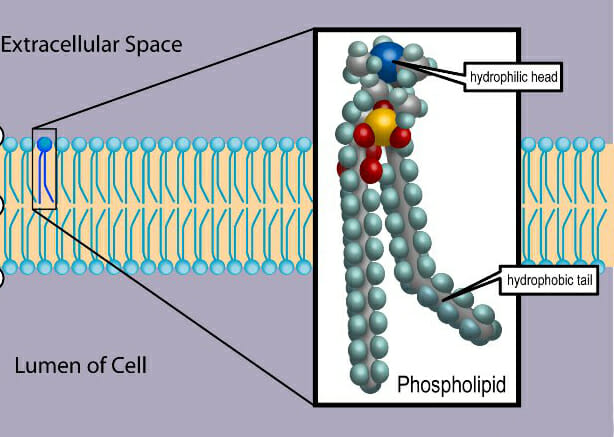
As seen in the image, an unsaturated fatty acid has a kink in its structure. This is an important feature that affects the fluidity, tensile strength and permeability of the membrane.
Proteins
In addition, the membrane has three types of proteins. Integral membrane proteins span the entire membrane, usually with alpha-helices forming the transmembrane region. These proteins form channels and pores that allow the movement of large or polar molecules across the hydrophobic segment of membrane.
In addition, there could be proteins embedded in a single leaflet of the membrane. These proteins are often used in signaling cascades, and can act as carrier molecules, transducing a signal from one segment of the membrane and relaying it to another region. These membranes are called peripheral membrane proteins. Finally, some proteins are very lightly attached to the membrane, with only a short lipid tail inserted into the hydrophobic region.
The proteins of the membrane can be covalently attached to carbohydrates and form glycoproteins. These can interact with water molecules and stabilize the membrane, as well as serve as important tools for intercellular communication. They form receptors for hormones and neurotransmitters.
The other important role for glycoproteins is to create a sort of ‘cell signature’. When immune cells recognize these glycoproteins, they are able to distinguish the cells of the body from those of pathogens. For instance, blood grouping into A, B and O types, depends on the kind of glycoprotein present on the surface of red blood cells.
Cholesterol
The presence of cholesterol in the phospholipid layer allows the membrane to retain its permeability and integrity even in varying temperatures. It appears to be inserted in the middle of phospholipids Cholesterol prevents the compaction of the hydrophobic tails at low temperatures as well as the expansion of the membrane under heat. This way, small molecules like carbon dioxide and oxygen can always move freely across the membrane, while the cell retains its selective permeability for larger molecules.
Other Models for Membrane Structure
The fluid mosaic model was refined in the early 1980s, by two scientists called Mouritsen and Bloom to create the ‘mattress model’ for membrane structure. They demonstrated the fact that while earlier experiments had suggested that the entire membrane is fluid and allows free diffusion of proteins, there are in fact, subdomains within each membrane.
For instance, when a transmembrane protein has a hydrophobic region that is slightly longer than the average width of a cell membrane, the lipid bilayer deforms to accommodate this protein. If there are multiple proteins whose hydrophobic stretches do not exactly match membrane width, the lipid bilayer would end up looking like a mattress, with interspersed thicker and thinner regions.
The thicker regions would likely create a slope that lets proteins slide ‘downward’, leading to protein aggregation in some regions. Similarly, these deformations may result in the accumulation of specific lipids around those proteins. These observations tied in with earlier experimental data suggesting the presence of lipid rafts, and the preferential association of proteins with lipids supported the ‘mattress model’.
Modern models of membrane structure also take into account the effect of lipid composition. Cell membranes are formed of many hundreds of phospholipids and each of them is made of different fatty acid side chains. These fatty acids can be of different length and contain varying degrees of saturation. There are also thermodynamic considerations in studying membrane properties since, even at physiological temperatures, the thickness of cell membranes and the distribution of different lipids can change.
Finally, membranes also have structures called lipid rafts that consist of special lipids, and cholesterol and proteins attached to the membrane through glycolipids. Lipid rafts are important subdomains, especially for signal transduction.
Related Biology Terms
- Amphipathic Molecules – Molecules containing polar hydrophilic regions and non-polar hydrophobic regions.
- Antigen – Any molecule capable of producing an immune response.
- Signal Transduction – Transmission of information, in the form of electrical or chemical signals, from the exterior of the cell to the interior.
- Sphingolipids – Fatty acid derivatives of a molecule called sphingosine. Often seen in membrane lipid rafts.
- Thermodynamics – The branch of science dealing with the relationship between heat and fundamental quantities like energy, work, entropy and temperature.
Quiz
1. Which of these statements about the structure of membranes is true?
A. Made primarily of cholesterol molecules
B. Glycoproteins on the cell surface are necessary for immune recognition
C. Lipid rafts were predicted by early models of cell membrane structure
D. All of the above
2. Which of these are features of the fluid mosaic model of cell membranes?
A. Lipid bilayer formed by amphipathic phospholipid molecules
B. A mosaic formed by proteins, cholesterol, and other membrane components
C. The ability for lateral diffusion of membrane components
D. All of the above
3. Which of these ideas represent a refinement of the fluid mosaic model?
A. Differing thickness of the cell membrane in different regions, depending on integral membrane protein composition
B. Presence of lipid rafts for signal transduction
C. Melting of lipids at physiological temperatures and changes to lipid composition in different subdomains of the membrane
D. All of the above