Common Descent Definition
Common descent is a term within evolutionary biology which refers to the common ancestry of a particular group of organisms. The process of common decent involves the formation of new species from an ancestral population. When a recent common ancestor is shared between two organisms, they are said to be closely related. In contrast, common descent can also be traced back to a universal common ancestor of all living organisms using molecular genetic methods. Such evolution from a universal common ancestor is thought to have involved several speciation events as a result of natural selection and other processes, such as geographical separation.
Theory of Common Descent
The theory of common descent states that all living organisms are descendants of a single ancestor. Thus, the Theory of Common Descent helps to explain why species living in different geographical regions exhibit different traits, some traits are highly conserved among broad animal classifications (e.g., vertebrates or tetrapods), seemly different species (e.g., birds and reptiles) share inherited physical and genetic traits, and successfully adapted organisms typically produce more offspring. While the Theory of Common Descent is primarily derived from the physical observation of various phenotypes (e.g., size, colour, beak shape, embryological development, etc.), modern advances in genetics and associated molecular techniques have been able to demonstrate that the process by which DNA is eventually translated into proteins is maintained among all lifeforms. Small changes in DNA between organisms have revealed a shared ancestry as well as insight into important changes that resulted in various speciation events. Phylogenetic trees and cladograms are often used to hypothesize the evolution of various organisms and shared common descent.
Examples of Common Descent
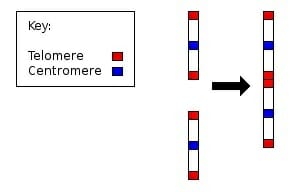
Human Chromosome 2
Compelling evidence of the shared common ancestry of humans with the great apes is the fusion event which occurred when two chromosomes common in apes fused to form chromosome 2 in humans (as illustrated below). This resulted in humans having only 23 pairs of chromosomes, while all other hominidae have 24 pairs. The great apes (e.g., chimpanzees, gorillas, and orangutans) have two chromosomes with almost identical DNA sequences as that found in chromosome 2 of humans. Further evidence of such a fusion event is the residual presence of telomeres and a centromere, which indicate that the genetic information was historically found on two separate chromosomes.
Endogenous Retroviruses
Endogenous retroviruses are residual DNA sequences found in the genomes of virtually all living organisms as a result of ancient viral infections. Since the retroviral sequences are incorporated into the DNA of the host organism, such sequences are inherited in the offspring. Since such infections are random events, as is the location in which the viral genome is inserted within the genome, the identification of the same retroviral sequences in multiple species is indicative of a shared ancestry. Such analyses of endogenous retroviruses often reveal speciation events (e.g., feline endogenous retroviruses reveal the separation between large and small cat species) and how closely related two species may be, as observed in the shared endogenous retroviruses between humans and other primate species.
The Presence of Atavisms
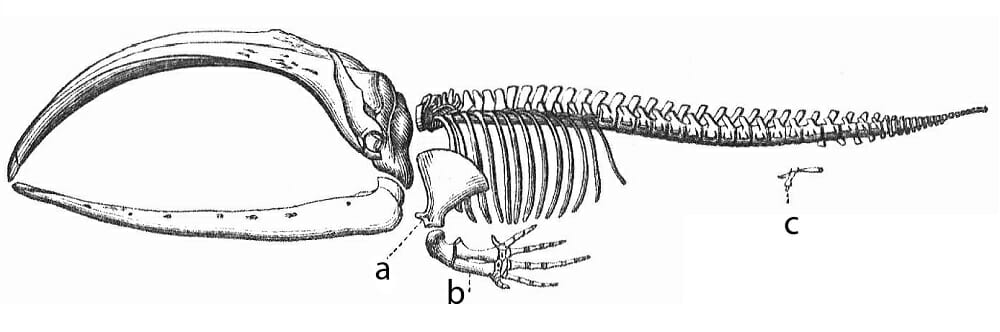
Atavisms are the appearance of a lost trait observed in an ancestral species that is not observed in more recent ancestors. Atavisms are an example of common descent as they provide evidence of the phenotypical or vestigial features that are often retained throughout evolution. Some examples, include the appearance of hind limbs in whales as evidence of a terrestrial ancestor, teeth exhibited by chickens, additional toes observed in modern horse species, and the back flippers of bottlenose dolphins. Atavisms tend to arise because the ancestral genes are not deleted from the genome, but rather silenced and then reactivated in later offspring.
Vestigial Structures
Similar to atavisms, vestigial structures are structures that are homologous to those found in ancestral species, but have become underdeveloped, non-functional, or degenerated in more recently evolved organisms. Such structures provide evidence of adaptations to a new environment, in which the ancestral organ or limb is no longer required, or has been modified to better suit a new purpose. There are an abundance of examples of vestigial structures observed in nature. Some examples include, the hind limbs and pelvic girdle observed in whales (as shown below) and snakes, non-functional wings exhibited by some insect species, non-functional wings of flightless bird species (e.g., ostriches), abdominal segments of barnacles, and the embryonic limb buds exhibited by several species lacking hind limbs (e.g., dolphins).
Pentadactyl Limbs
The presence of pentadactyl limbs is an example of a homologous trait exhibited by all tetrapods and is highly conserved throughout evolution, despite some modifications. Such limbs are first observed in the evolution from fishes to amphibians and consists of a single proximal, two distal, carpal, five metacarpal, and phalange bones. Although the overarching structure of the pentadactyl limbs is similar, various modifications have been made throughout evolution as adaptations to specific environments or lifestyles. Some examples include the modified pentadactyl wings of bats, the elongated forearms of primate species, the flippers of dolphin and whale species, and the modified digits of horses to form a hoof.
Fossil Evidence
The fossilized remains of various organisms combined with modern dating methods provides some of the most convincing evidence of common descent and evolutionary history. Fossilization occurs when the bones of a decaying animal become porous and the mineral salts in the surrounding earth infiltrate the bones, converting them to stone. Other methods include preservation in ice, imprinted remains (e.g., plants or footprints), tree resin, and peat. Since fossils are found in sedimentary rock, which is formed by layers of silt and mud, each layer corresponds to a specific geological period which can be dated. Thus, the extinction, evolution, and emergence of various species can be observed throughout history using the fossil record. Many extinct species are also observed in the fossil record, such as dinosaurs.
Biogeography
Biogeography presents compelling evidence of common descent by showcasing speciation and novel traits through adaptations to environmental pressures. One of the most famous examples is that of island biogeography and Charles Darwin’s observations of the beaks of finches residing on the Galapagos Islands. In these finches, the beaks had been adapted for the specific vegetation found on the island, resulting in deviation from the ancestral finches found on the mainland. Long term effects of geographical separation are also observed with the evolution of new species that are not found elsewhere in the world. An example of this is the presence of marsupial species on the continent of Australia, and the emergence of polar bears as a result of geographical isolation in the arctic.
Quiz
1. The presence of a tail during the early stages of human embryological development is an example of:
A. Sequence homology
B. Biogeography
C. Activation of chromosome 2
D. Vestigial structure
2. Recently, a bottlenose dolphin with four fins (two in the front and two in the back rather than only two in the front) was identified. This is an example of:
A. Convergent evolution
B. Pentadactyl limbs
C. Activation of chromosome 2
D. An atavism
References
- Costa J. (2009). The Darwinian Revelation: Tracing the Origin and Evolution of an Idea. BioScience. 59(10): 886-894.
- Koonin E and Wolf Y. (2010). The common ancestry of life. Biology Direct. 5:64.
- Wall J. (2013). Great Ape Genomics. ILAR J. 54(2): 82-90.