Crossing Over Definition
Crossing over is the exchange of genetic material between non-sister chromatids of homologous chromosomes during meiosis, which results in new allelic combinations in the daughter cells.
Each diploid cell contains two copies of every chromosome, one derived from the maternal gamete and the other from the paternal gamete. These pairs of chromosomes, each derived from one parent, are called homologous chromosomes. When diploid organisms undergo sexual reproduction, they first produce haploid gametes through meiosis. During prophase I of meiosis, homologous chromosome align with each other and exchange genetic material, so that some of the resultant chromosomes are recombinants – containing a mixture of genes derived from the maternal as well as the paternal chromosomes.
Homologous Chromosomes
Most organisms that undergo sexual reproduction contain two types of cells in their body – haploid and diploid cells. Diploid cells are seen in most parts of the body and contain two copies of every chromosome. Therefore, they contain two genes to determine the sequence of nearly every RNA or protein. Haploid cells are usually male or female gametes formed as a result of meiosis and are seen in sexual organs. They contain only one complete set of chromosomes and are designed to fuse with another haploid gamete to produce a diploid zygote – with one paternal and one maternal set of chromosomes. Though they code for the same genes, their DNA sequences can vary slightly from one another. For instance, the paternal chromosome may contain the genetic information to result in antigen A being present on red blood cells, while the maternal chromosome may code for antigen B. These two antigens are similar in function, but differ from each other marginally in their amino acid sequence.
All the cells of an adult organism are derived from the initial zygote and are therefore genetically identical to each other. When this adult prepares for reproduction, some specialized cells undergo meiosis, producing four haploid daughter cells. These daughter cells can then undergo maturation to form male or female gametes.
During meiosis, chromosomes derived from the maternal and paternal gamete can assort independently from one another. In other words, chromosomes from the mother or the father can be chosen randomly for each gamete. A human sperm can contain chromosomes 1, 3, 5, 7, 9, 21 and 23 from his female parent, and the rest can arise from the male parent. Similarly, an egg can have only two chromosomes from the female parent and the rest from the male parent.
In addition, each of these chromosomes could be a recombinant mixture of genes arising from the two parents. One of the four haploid daughter cells arising from meiosis could have a chromosome with 80% of its sequence from the male parent and the remaining 20% from the female parent. Correspondingly, there will be another daughter cell with the reverse ratio. This occurs through crossing over, when replicated homologous chromosomes are placed close to one another and exchange segments with each other.
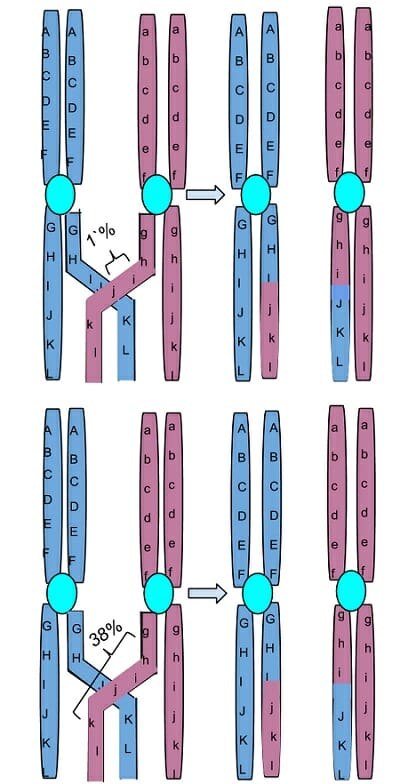
The image is a representation of one set of homologous chromosomes, with genes being represented by different letters of the alphabet. The genes on one chromosome are shown using capital letters and that on the homologous pair using small letters. Two of the four daughter cells formed after this crossing over event have a recombinant chromosome that is neither completely derived from the mother nor the father. The image also demonstrates that genes that are in close physical proximity to one another on the chromosome are likely to be inherited together, while those that are farther away might get independently assorted during meiosis.
Linkage Maps
Linkage maps are estimates of the distance between two genetic loci, based on the frequency of recombination. Gene loci that are far away from each other on the same chromosome, or those placed on different chromosomes have a 50% chance of being inherited together and a 50% probability of being assorted independently. Based on how frequently they are inherited together, linkage maps can be formed, where the position of these genes is shown in relation to known genetic markers. For instance, if a genetic marker on the X chromosome is frequently inherited along with a particular allele for male pattern baldness, they are said to be part of the same linkage group. Prior to high throughput DNA sequencing, linkage maps were the primary tools to understand which genes were present on a particular chromosome.
Prophase I
Meiosis occurs in two stages – meiosis I and II. Meiosis I, also known as reduction division, is the series of events that results in the formation of two haploid daughter cells. At the end of reduction division, the number of chromosomes is halved and each of the daughter cells has only one complete set of duplicated chromosomes.
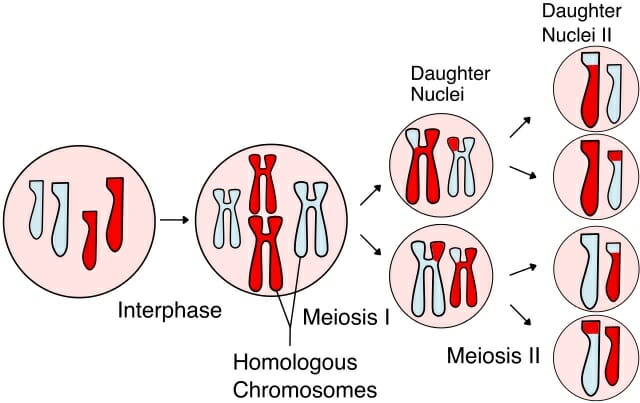
During meiosis I, particularly prophase I, a number of events occur, making it one of the longest phases in meiosis. It marks the beginning of chromosome condensation where duplicated chromosomes with attached sister chromatids, are initially seen as long thin threads. As condensation proceeds, homologous chromosomes are brought together because of the similarity in structure and centromere position. A protein structure called the synaptonemal complex also plays an important role. At this point, chromosomes are anchored to the nuclear envelope. Now, recombination occurs between non-sister chromatids of homologous chromosomes. This is observed microscopically as a crossing over event between bivalent chromosomes (a pair of two chromosomes) with a tetrad structure (their duplicated sister chromatids are also visible). Towards the end of prophase I, homologous chromosomes now appear to ‘repel’ each other. The nuclear envelope is no longer clearly visible and the cell then moves on to metaphase and anaphase to complete the first stage of meiosis.
Mechanism of Crossing Over
On a molecular level, crossing over begins with a double strand break in one of the DNA molecules. This double strand break can occur naturally through agents like radiation or carcinogens, or through the action of specific proteins. Subsequently, exonucleases, enzymes that remove nucleotides from the 5’ end of DNA, act on this break and remove short stretches of nucleotides in the 5’ -> 3’ orientation from both the strands. This leads to two hanging single-stranded regions that get coated with proteins that catalyze recombination, also known as recombinases. These enzymes catalyze the invasion of single strand regions into sequences that are suitable for base pairing. The close proximity of non-sister chromatids during prophase I, allows this single-stranded region to use the sequence on the homologous chromosome. The first invading strand behaves like a primer and synthesizes a double stranded region for itself using one strand of its non-sister chromatid as a template. This leads to its complementary strand getting displaced and base pairing with the second single stranded region that was initially generated by the exonuclease. Ultimately, this results in two strands being exchanged with the formation of a cross-like structure called the Holliday junction. This is named after the scientist who first proposed that such a junction could explain both crossing over and another phenomenon called gene conversion where a heterozygous gene locus becomes homozygous during cell division. Holliday junctions can also be seen microscopically as ‘chiasma’ towards the end of prophase I, which continue to be visible till the end of anaphase I. Holliday junctions are stabilized and resolved through proteins that modulate genomic manipulation which are known as MSH4 and MSH5.
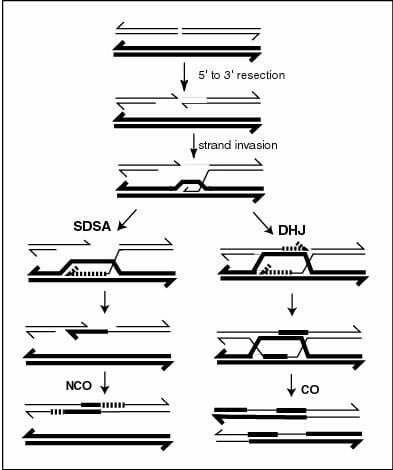
In the image, the events after strand invasion that lead to crossing over and Holliday junction formation are given on the right. Non-crossing over events, including gene conversion are depicted on the left.
Functions of Crossing Over
Organisms that divide only asexually without the chance of such recombination suffer from a condition called Muller’s Ratchet. That is, each generation of that species contains at least as many genetic mutations as the previous generation, if not more. In other words, when all the progeny are genetically identical to one another, there is no scope for genetic errors to be corrected, or for new and beneficial combinations to arise.
Crossing over increases the variability of a population and prevents the accumulation of deleterious combinations of alleles, while also allowing some parental combinations to be passed on to the offspring. This way, there is a balance between maintaining potentially useful allelic combinations as well as providing the opportunity for variation and change.
Related Biology Terms
- Cytokinesis – Division of the contents of the cytoplasm into two daughter cells at the end of mitosis or meiosis.
- Karyokinesis – Division of the contents of a nucleus during cell division.
- Ploidy – The number of sets of chromosomes in a cell. While haploid (one set) and diploid (two sets) are most common, some organisms can contain triploid, or even tetraploid cells.
- Zygote – A diploid cell formed as a result of the fusion of two haploid gametes.
Quiz
1. When does crossing over occur?
A. Mitosis
B. Meiosis II
C. Prophase I
D. Metaphase I
2. Which of these statements about crossing over is NOT true?
A. Is necessary for the formation of diploid gametes
B. Double strand breaks occur through the action of ribonucleases
C. Leaves organism vulnerable to Muller’s Ratchet
D. All of the above
3. Which of these numbers represents the chromosome number in somatic cells and gametes?
A. Somatic cells are haploid, gametes are diploid
B. Somatic cells are diploid, gametes are haploid
C. All cells are derived from a diploid zygote and therefore are also diploid
D. All cells are haploid