Definition
The dinoflagellate is a single-celled aquatic organism with two flagella. It is known to cause bioluminescence in the ocean. Dinoflagellates are found in fresh and saltwater and, in large numbers, can produce dangerous levels of neurotoxic chemicals. These organisms – halfway between plant and animal – convert sunlight as well as consumed organic matter to energy. Dinoflagellates are food chain producers and, as a component of plankton, provide an important source of nutrition for aquatic primary consumers.
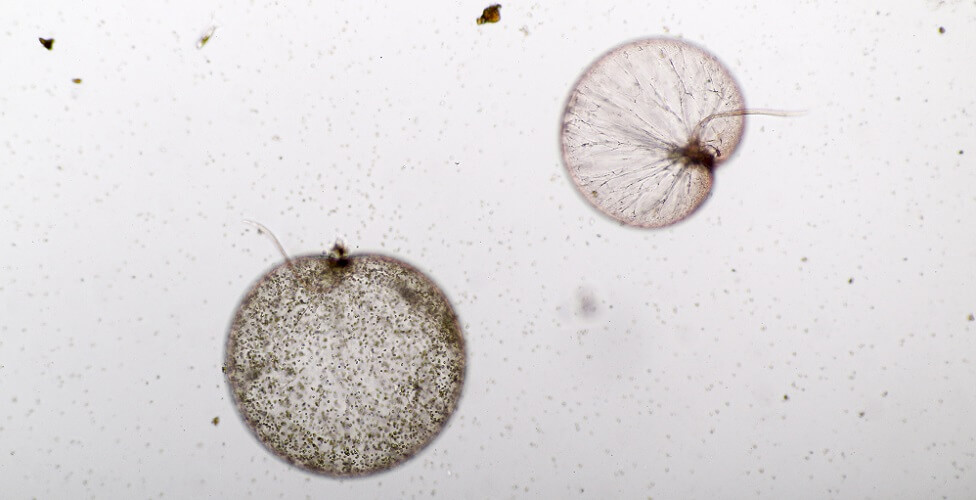
Dinoflagellate Anatomy
Dinoflagellate anatomy is complex, even though these are unicellular eukaryotes. As part of the earliest forms of life, they straddle the border between plant and animal.
The name dinoflagellate is a combination of the Greek words for whirling (dinos) and whip (flagellum). The suffix describes the two flagella. Little variety is found in shape; however, the dinoflagellate Ceratium genus is usually elongated (fusiform) with horns.
Basic dinoflagellate morphology is an armored (thecate) or unarmoured (athecate) cell with a transverse and longitudinal flagellum.
The transverse flagellum is wave-like and wraps around the cell body. It pulses to the left, moving the dinoflagellate forward and turning it. The longitudinal flagellum hangs below the cell and beats more slowly.

Thecate dinoflagellate cell walls are composed of cellulose plates that surround the amphiesma. The amphiesma consists of the cell membrane, alveolae (structural vesicles), and multiple organelles. In thecates, the cell’s walls extend from these vesicles and overlap to produce an armored covering. Some dinoflagellate species have another layer of cellulose under the alveolae called the dino-pellicle.
Inside the membrane, several sacs or vacuoles are formed from pusule ducts. As yet, we do not know their role, although they are more complex in heterotrophic dinoflagellate species that do not get any energy from the sun. They, therefore, probably play a role in digestion.
Organelles inside the dinoflagellate organism are found in large numbers. They include mitochondria, a nucleus, and secretory cysts.
Very few dinoflagellates use only sunlight to produce energy. Most are mixotrophic rather than purely photosynthetic (homotrophic) and digest particles located in their watery environment. Homotrophic phototropic and mixotrophic dinoflagellates contain plastids that manufacture and store food produced from photosynthesis. This food is stored in the form of amino acids, starch, and lipids.
One dinoflagellate class – Noctiluciphyceae – is grouped because of its ability to steal plastids (kleptochloroplasts) from surrounding algae. Chloroplasts are a plastid subtype. Inside the plastids of some dinoflagellates are compartments that help the cell to produce a carbon source that can be converted to starch.
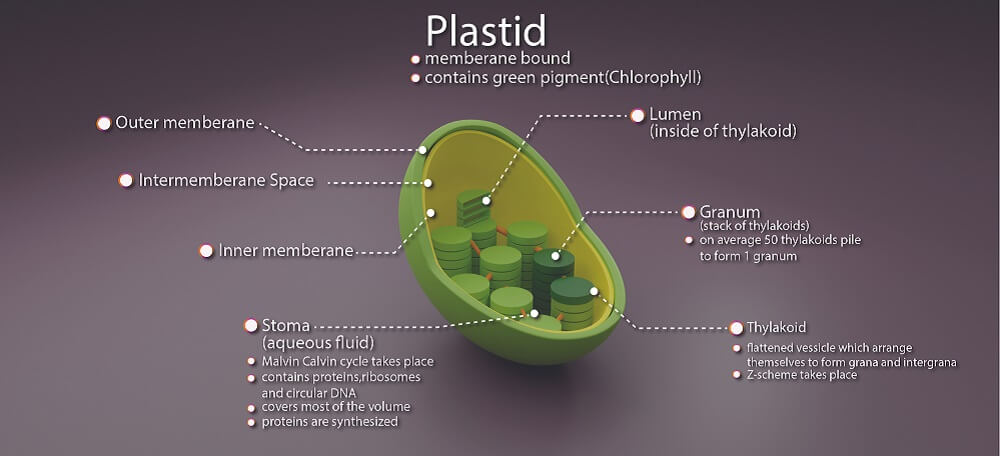
Dinoflagellates consume nutrients by way of osmosis and phagocytosis. Phagocytosis requires the cell to be athecate – without armor. Athecate cells have a cytosome or cell mouth. Some cells feed on other unicellular organisms by puncturing their membranes and sucking out the contents with a hollow tube called a peduncle. Armored dinoflagellates are limited in how much they can consume; they sometimes surround food sources with a membrane filled with digestive enzymes called a pallium.
The dinoflagellate stigma or eyespot may play a role in light reception. Stigmas seem to aid movement – the many dinoflagellate eyespots help it move toward light (positive phototaxis) and may control when luminescence occurs.
Many dinoflagellate genera can produce light – they are bioluminescent. This occurs in scintillons dotted around the edge of the vacuoles. Scintillons are sacs filled with the enzymes luciferase and luciferin. These two enzymes are kept separate and only create light when they come into contact with each other. When they mix, the reaction produces a blueish light that either glows gently or pulses strongly. This only happens at night when the eyespots are unable to detect light. Dinoflagellate circadian rhythm seems not to be controlled by light levels.
Dinoflagellate Life Cycle
Eighty percent of dinoflagellates are found in marine environments and the rest in freshwater habitats. Most are phototropic and convert sunlight into energy as well as gain energy from other organisms as heterotrophs. This combination of phototrophic and heterotrophic characteristics is referred to as mixotrophic. Most dinoflagellates are mixotrophic.
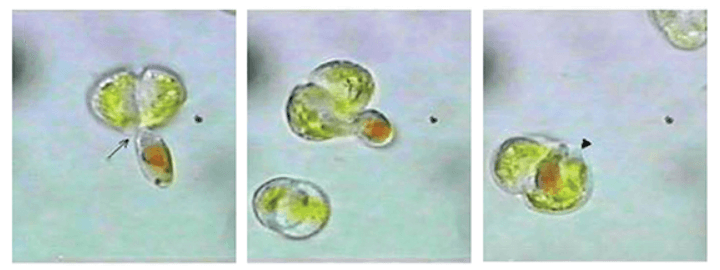
Dinoflagellates are haploid during the majority of their life cycle. This means cells contain a single set of chromosomes. They multiply via cell division (asexual fission); however, some species reproduce when they become gametes (sex cells) that fuse to form planozygotes. This usually occurs when the environment is hostile, as planocytes become walled, dormant cysts called hypnozygotes until conditions improve.
Interestingly, the DNA of fused dinoflagellates undergoes a process similar to crossing over in more complex multicellular organisms. During nuclear cyclosis, the two single chromosomes are mixed. They later divide to form two haploid cells with mixed alleles. Mixing alleles causes increased diversity.
Dinoflagellate cysts travel along currents or are consumed by other animals and are, therefore, easily spread. This feature has made dinoflagellates extremely successful.
Dinoflagellate Genus
To note down the entirety of the dinoflagellate genus would take some time. There are well over three thousand known dinoflagellate species split into various genera. Even more confusing is this genus’ classification. Although a eukaryote, dinoflagellates can be found in both botanical and zoological codes of nomenclature throughout history.
More recently, dinoflagellate evolution has been looked at in detail. Our increased knowledge of molecular structures has produced very different classification groups compared to previous attempts.

The older dinoflagellate taxonomy is flawed. Classification according to shape and structure (morphology) is becoming outdated as gene analysis of dinoflagellate DNA moves forward.
Until DNA sequencing fully uncovers the cladistics (the evolution of shared characteristics) of dinoflagellates, outdated classification methods still apply. Most differ according to source and only look at cellular structure and lifestyle. This type of classification is called linear taxonomy.
The Euteleost Tree of Life project is gaining recognition in scientific circles. This non-molecular phylogenetic (ancestral history) searchable chart shows us dinoflagellate classification from Domain to Superclass. From Superclass on, classes, orders, families, genera, and species remain extremely similar to older methods. The dinoflagellate phylogenetic tree runs as follows:
- Domain: Eukaryotes
- Kingdom: Chromalveolata
- Supergroup or Clade 1: SAR
- Clade 2: TSAR
- Clade 3 or Infrakingdom: Alveolata
- Phylum: Miozoa
- Superclass: Dinoflagellata
- Class:
- Dinophyceae
- Ellobiophyceae
- Noctiluciphyceae
- Oxyrrhidophyceae
- Syndiniophyceae
- Order: for example, Gymnodiniales of the class dinophyceae that does not have an armored (thecate) exterior.
- Family: for example, Oxyrrhinaceae of the order Oxyrrhinales and the class Oxyrrhidophyceae.
- Genus: for example, Alexandrium. Produces highly-toxic algal blooms.
- Species: for example, Dicroerisma psilonereiella of the genus Dicroerisma, family Actiniscaceae, Order Actiniscales, and class Dinophyceae. A marine species first found in Canada.
The chromalveolata Kingdom theory is currently under much discussion – it groups non-plants into organisms that create energy using photosynthesis. This characteristic has been lost in some dinoflagellate genera. However, even though this feature is no longer expressed, the genetic code for photosynthesis can still be found in non-phototropic dinoflagellate DNA.
TSAR refers to a unicellular eukaryotic supergroup composed of stramenopiles (two flagella), alveolates, and Rhizaria (amoeba-like). At a later evolutionary point, the telonemid taxon (formerly called the Protist Kingdom) was added (TSAR). Each attached group (clade) shares a common ancestor and is created according to an evolutionary timeline.
Dinoflagellate Toxin
The dinoflagellate Alexandrium genus produces red-colored toxins that accumulate in shellfish but do not kill them. When humans consume contaminated shellfish – even if cooked at high temperatures – they may suffer from paralytic shellfish poisoning.

The primary neurotoxin produced is saxitoxin. Symptoms appear rapidly and include nausea, abdominal cramps, numbness in the mouth, pins and needles in the extremities, shortness of breath, an inability to balance, and slurring speech. Paralytic shellfish poisoning can be fatal. It is known to cause the death of large populations of marine mammals.
The dinoflagellate Amphidinium is another potentially toxic genus, although rarely fatal. As it stays close to the sand even at night (unlike most types of dinoflagellate), its dormant cysts may contaminate aquarium sand sold for marine fish displays.
Dinoflagellate Bioluminescence
Dinoflagellate luciferin is a compound that emits light when oxidized. This oxidation reaction is only possible in the presence of dinoflagellate luciferase – another enzyme.
The luciferin-luciferase reaction occurs in the cell scintillons and was first observed in the laboratory in fireflies. However, this mechanism differs from that of the dinoflagellates; dinoflagellate luminescence needs no adenosine trisphosphate. Firefly luminescence does.

To initiate the chemical reaction, acidity levels in the cell vacuole need to be approximately pH 6.3. This causes luciferin release from luciferin binding protein. Dinoflagellate luciferinase enzymes can now bind to the released luciferin and, in the presence of oxygen, an intermediate molecule is produced. It is this highly-reactive intermediate that oxidizes the bound luciferin-luciferinase complex and emits light. The process is called proton-stimulated luminescence.
There are two types of dinoflagellate bioluminescence: flashing light emission and glow light emission. Flashing is activated by changes in vacuole acidity and the light produced is bright and pulsating. Glow is a constant light that is not at all intense.

Blue light travels furthest through water and dinoflagellates are the main eukaryotes that produce bioluminescence in the ocean. Marine phosphorescence was recorded as early as 500BC by the Greeks.
Why dinoflagellates produce bioluminescence is not yet known. Theories include the reaction’s prevention of the toxic effects of oxygen. They also mention its usefulness as an alarm signal during dinoflagellate attack by marine consumers and first-order predators . This alert might attract second-order predators that consume dinoflagellate feeders.