Selective Permeability Definition
Selective permeability is a property of cellular membranes that only allows certain molecules to enter or exit the cell. This is important for the cell to maintain its internal order irrespective of the changes to the environment. For example, water, ions, glucose and carbon dioxide may need to be imported or exported from the cell depending on its metabolic activity. Similarly, signaling molecules may need to enter the cell and proteins may need to be released into the extracellular matrix. The presence of a selectively permeable membrane allows the cell to exercise control over the quantum, timing and rate of movement of these molecules.
Movement across a selectively permeable membrane can occur actively or passively. For example, water molecules can move passively through small pores on the membrane. Similarly, carbon dioxide released as a byproduct of respiration quickly diffuses out of the cell. Some molecules are actively transported. For example, cells in the kidney expend energy to reabsorb all the glucose, amino acids and vitamins from the glomerular filtrate even against the concentration gradient. Failure of this process leads to the presence of glucose or the byproducts of protein metabolism in urine; a tell tale sign of diabetes.
Structure of Selectively Permeable Membranes
Cell membranes are not easily visualized using light microscopes. Therefore, hypotheses about their existence only arose in the late 19th century, nearly two hundred years after the first cells has been observed. At various points, different models have attempted to explain how the structure of the membrane supports its function. Initially, the membrane was supposed to be a simple lipid layer demarcating the cytosol from the extracellular region. Afterwards, models included semipermeable gel-like regions in a lipid sea to explain the movement of water but not charged particles. Thereafter, the presence of pores was proposed, allowing small molecules to move freely.
Currently the cell membrane is said to be made of a selectively permeable phospholipid bilayer whose hydrophillic domains face the aqueous environments inside and outside the cell, and hydrophobic domains face each other to form a bilayer. This lipid bilayer is punctuated by cholesterol molecules, glycolipids, and proteins that are either anchored or traverse the entire membrane. These proteins form channels, pores or gates to maintain selective permeability of ions, signaling molecules and macromolecules based on the requirements of the cell.
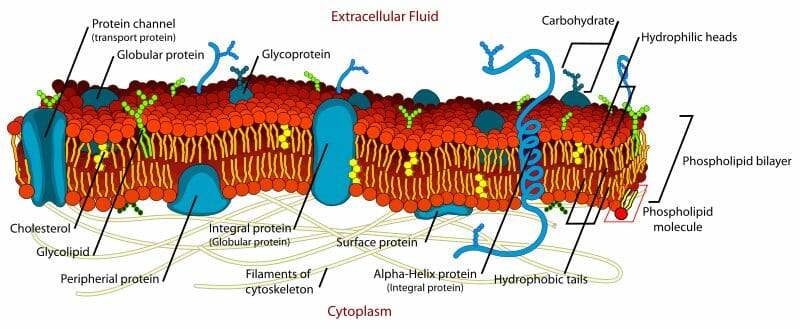
The Nuclear membrane has a structure different from all the other membranes of the cell. It has nuclear pore complexes – basket-like multiprotein complexes that are freely permeable to water but stringently mediate nuclear transport of macromolecules. Importins and exportins are two classes of proteins that are actively involved in nuclear transport. Both are energy intensive and each transport event involves the hydrolysis of a high energy phosphate bond on a guanosine triphosphate. The directionality of the movement also needs the presence of a small molecule called Ran, that has a differential affinity for its substrates based on whether it is bound to GTP or GDP.
Function of Selective Permeability
Selective permeability is crucial for creating a distinctly different environment inside the cell as compared to the extracellular matrix. It is equally relevant in maintaining the integrity of various organelles inside the cell. Each organelle is a small compartment with a specialized function, requiring optimal concentrations of proteins, small molecules and ions. For instance, cellular respiration inside a mitochondrion requires that the proteins that aid this process be selectively imported into the organelle, and its internal chemistry should remain unaffected by the other metabolic processes of the cytoplasm. Similarly, after a neuron transmits an electrochemical signal, it needs to recover and return to its resting potential to enable the next round of excitatory activity. The same happens in every cardiac muscle cell each time the heart beats. These rapid and large scale changes in electrochemical properties of these cells are necessary for their function and need the presence of a membrane that is selectively permeable.
Selectively permeability of membranes is particularly important for transport across the nuclear membrane in eukaryotic cells. Proteins, nucleic acids, and nucleotides involved in transcription must be selectively and efficiently transported into the nucleus and the products of transcription must be exported in a timely manner. The nucleus has a distinct microenvironment compared to the cytoplasm and active transport mechanisms are at work to maintain this distinction.
Proteins Mediating Selective Permeability
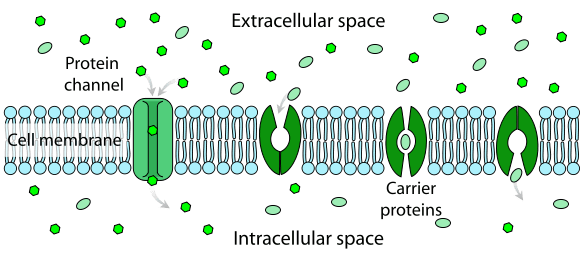
Selective permeability is mediated by special proteins that traverse the cellular membrane. They are involved in the movement of ions and small molecules as well as large polymers such as RNA and proteins. This movement can be passive or active – with or without the expenditure of energy.
For instance, ions are transported across selectively permeable membranes through channels and pumps. While channels are for passive transport, ion pumps mediate primary active transport against a concentration gradient, with the hydrolysis of a high-energy phosphate bond.
Active transport can also be coupled with the movement of another molecule. This can either be through a symporter protein – where two molecules are transported in the same direction – or antiporter protein – where molecules are shunted in opposite directions. The principle in both cases is the same – the potential energy stored in an electrochemical gradient is used to drive the transport of another molecule.
Active And Passive Transport Across Selectively Permeable Membranes
Passive transport is of two types – free diffusion or facilitated diffusion – and movement is always along a concentration gradient. Free diffusion is seen most often in the movement of uncharged molecules such as carbon dioxide or ethanol across the cell membrane, without the involvement of any other molecules.
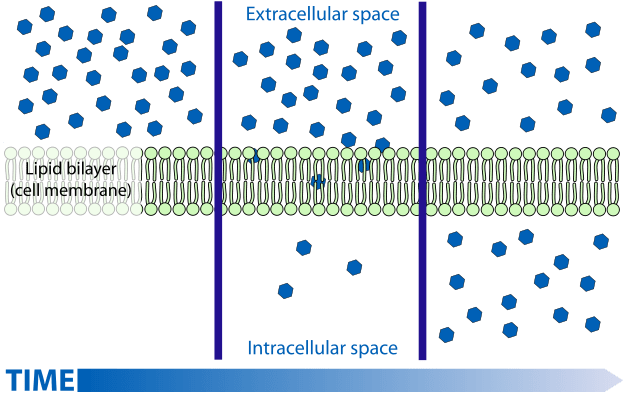
Facilitated diffusion requires the presence of another molecule, usually a protein, that acts as a carrier and helps the substrate cross the cell membrane. Carrier proteins bind to the substrate on one side of the membrane and change conformation to release the substrate on the other side. Classic examples of facilitated diffusion are the movement of oxygen through binding to haemoglobin or the transport of water through minute pores formed by aquaporins.
The diffusion of water can be observed at the macroscopic level as well. For example, when seeds swell after being soaked in water, we are seeing the overall effect of water entering the cell. Similarly, fruits left in a dry environment, such as a refrigerator, shrivel and shrink as they lose water. Many organisms, including humans, have a waxy coating over their skin to minimize water loss from their cells in a dry environment.
Transmembrane transport can also be accomplished actively, with the expenditure of energy. Active transport involves the hydrolysis of the terminal phosphate group in ATP or GTP to power the movement of molecules against their concentration gradient. For example, in most cells, there is a large excess of sodium ions in the extracellular environment along with an excess of potassium ions inside the cell. This is achieved by a transmembrane enzyme called the Na+/K+ ATPase, that catalyzes the movement of three Na+ ions outside the cell along with the import of two K+ ions. For each such transport cycle, the enzyme uses the energy released from the conversion of one molecule of ATP into ADP. This is called primary active transport, where the movement is directly coupled to the hydrolysis of a high-energy phosphate bond. A similar process is used to pump protons against their concentration gradient and this is a crucial part of both photosynthesis and cellular respiration.
Gradients of H+, Na+ and K+ ions are used to drive other processes, through secondary active transport, where differential electrochemical concentrations provide the driving force for other energy intensive processes, such as the transport of amino acids or glucose. For instance, absorption of glucose in the intestine is coupled to the transport of Na+ ions. This is an example of symport, where both the sodium ion and glucose molecule are imported into the cell. Sodium ions are also involved in the movement of another charged molecule – Ca2+. The sodium-calcium exchanger uses the movement of Na+ along its gradient to drive the counter transport of Ca2+. This is particularly important in the movement of calcium ions in large quantities such as in neurons, cardiac cells, and for the maintenance of low concentration of calcium in the mitochondria.
Related Biology Terms
- Electron Microscope – A microscope that uses a beam of electrons to illuminate the sample achieving extremely high magnification and resolution.
- Extracellular Matrix – Non-cellular component of tissues and organs composed of water, proteins and polysaccharides, which provides physical, biomechanical and biochemical support to cells.
- Hydrophile – A molecule that is attracted to water.
- Ran – A small protein that directs transport across the nuclear membrane based on its binding to guanosine dinucleotides and trinucleotides.
Quiz
1. Which of these proteins is involved in nuclear transport?
A. Importin
B. Exportin
C. RanGTP
D. All of the above
2. Which of these molecules diffuses freely through small pores on the cell membrane?
A. Glucose
B. ATP
C. Water
D. None of the above
3. Choose the molecule that does NOT need active transport to move across the cell membrane.
A. Sodium ions
B. Carbon dioxide
C. Amino acids
D. Potassium ions